|
Continued from Relative Length of the Intestinal Canal
In early embryonic stages the Intestinal Canal is
a straight tube; but, as its growth proceeds far more rapidly than that
of the body-cavity, it is necessarily thrown into folds or loops. Moreover,
since it is suspended from the vertebral column by the mesentery, or lining
of the body-cavity, its several folds are thereby connected with one another
in various ways, and their number and shape depend to a great extent upon
the space available in the cavity, as well as upon the shape, size, and
position of the stomach and neighbouring organs; but the various ways in
which the small intestine is stowed away in different birds exhibit types
so definite and constant that they cannot be considered accidental or meaning-less
features.
On the contrary, a somewhat exhaustive study of
its convolutions reveals their taxonomic value, and enabled A. Newton to
say that the Digestive System; taken in its entirety-that is to say, the
crop, glandular and muscular stomach, liver, gall-bladder with its ducts,
caeca, and the relative length and convolutions of the intestinal canal
- affords more diagnostic features than any other organic system - the
oseous excepted. Moreover, it has the great advantage that through reference
to the food we can in many cases account for the aberrant features of the
digestive organs displayed by birds otherwise closely allied. So much cannot
be said for characters furnished by PTERYLOSIS, and attempts to explain
taxonomically the important differences observable in the MUSCULAR
System have hitherto been futile because of the complex problems involved.
At any rate, we ought not to treat recent birds as if they were fossil
and had left us nothing but their bones, unless, indeed, the specimens
be skinned and all their other important characters thrown away.
It is hoped therefore that a brief general account
of the chief types of intestinal structure in birds may here have interest,
especially as, since the days of Macgillivray, who alone attempted it systematically,
this branch of Ornithology has been relatively neglected, perhaps from
the apparent but not real difficulty of studying these easily-putrefying
organs.
In a typical loop of the intestines of a bird we
distinguish between a descending and an ascending branch; both meet at
the distal end or apex of the loop, and this forms its turning-point. 'The
starting-point is the pylorus, the goal the cloaca.
Each loop is either closed or open. It is closed when both the descending
and the ascending branches are throughout the length of the loop closely
bound together by an extension of the mesentery and its vessels.
Of these vessels, as a rule, each principal loop receives one bigger branch
from the middle mesenteric artery. A loop is open when its two branches
are not closely connected by mesentery and vessels, the mesentery is wider,
and the two branches of the loop may receive another loop or intestinal
fold between them, the latter then resting upon the mesentery of the former
open loop.
'I'he duodenum is always a typically-closed loop.
Its first or descending branch lies, when viewed from the ventral side,
to the right of the second or ascending branch; both invariably enclose
the PANCREAS.
A loop which runs in the same way as the duodenum
may be termed right-handed,
and one running in the opposite way
is left-handed, i.e. its descending branch lies to the left of the
ascending branch, Again, if the intestine forms a number of (mostly closed)
loops, which run parallel with each other in the long axis of the body,
we term this arrangement orthocoelous,
or straight-gutted.
If, on the other hand, some of the loops form a
spiral, we distinguish this formation as cyclocoelous.
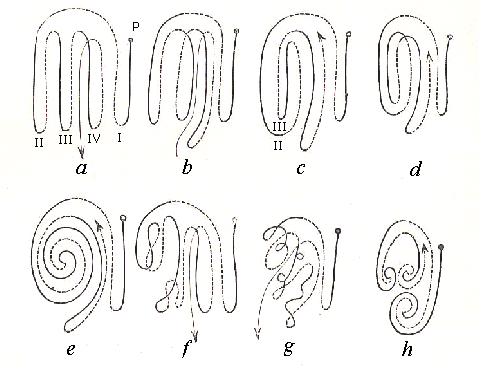
Diagram of a the principal
relative positions of the Intenstinal Loops when seen from the right side
a..... Isocoelous
b..... Anticoelous
c..... Anti-Pericoelous
d.....
Iso-Pericoelous
e..... Cyclocoelous
f and g.....
Plagiocoelous
h.....
Telogyrous
The descending branches of the loops
are marked by black lines, the ascending or returning branches are dotted.
The first and third loops in figure b are "right-handed," the second is "left-handed";
in figure c the second is "left-handed", the third "right-handed," etc. |
Of the orthocoelous type the following modifications
deserve especial remark with reference to the second and third loops; the
first, or duodenal, loop is invariably right-handed, and therefore needs
no further comment.
I. Isocoelous. - The second and
third, and, if present, also the fourth loop are all closed and left-handed.
The second is most dorsally situated, the third to the right of it, the
fourth to the right of the latter, between it and the duodenum. The ascending
branch of on loop runs side by side with the descending branch of the next
one.
II. Anticoelous. - The second and
third loops are closed and sharply alternating; the second is left-, the
third is right-handed; the second lies dorsally, consequently its ascending
branch runs side by side with that of the third.
III. Plagiocoelous - The second and
often more loops are doubled or turned over with the apices like a horseshoe,
giving the loops, which are generally open, an irregular or convoluted
appearance.
IV. Pericoelous - The second loop
is left-handed, open, and encloses the third which is generally straight
and closed. This formation is of especial interest, because it leads quite
gradually to...
V. The Cyclocoelous formation by the conversion
of the second and third loops into one left-handed spiral. Such a conversion
of the second and third loops into a spiral occurs in the Limicolae,
Laridae, and.Columbae.
Each of these families possesses some
genera in which the spiral is still represented by long, oval, concentric
turns, and even some genera which still exhibit the pericoelous type with
the two loops in question still separate, distinct, and more or less straight.
Not every spiral, however, is formed by the concentration
of two loops. In many instances a spiral is produced by one loop
being curled upon itself, its apex then forming the centre of the spiral.
To the apex is attached the diverticulum caecum vitelli; this shows that
this spiral is produced by the primitive fold of the embryonic mid-gut.
Such is the case in all the Passeres, and
since there are only three folds formed by the whole gut, the spiral represents
the middle or second fold; hence this arrangement may be distinguished
as mesogyrous.
The number of turns in such a spiral depends directly
upon the length of the intestine; while in the short-gutted Sylviae
the spiral is just indicated, there are in the Sparrow (with an intestinal
length of 21 centimmetres) 1½ direct and 1 retrograde turn, and
in Pinicola enucleator
(which possesses an intestine of 99 centimetres
in length) there are many direct turns.
It is clear that with an original number of only
four loops, the conversion of the two middle ones into a single spiral
will cause such birds as certain
Limicolae, Laridae, and Columbae
likewise
to assume the mesogyrous feature; but the position of the diverticle on
the original third loop, and the relations of these birds,
e.g. Charadrius
and
Sterna,
show that this mesogyrous formation has been brought
about in a way different from that of the Passeres.
Lastly the distal portion of any loop originally
straight may be coiled up into a spiral, while the rest of the loop remains
straight. This feature may be termed telogyrous. With the
duodenum this is very rare, it then invariably forms a right-handed spiral,
e.g. in Buceros, Ciconia, and Milvus; the duodenum is more
irregularly twisted in certain Pelargi and Accipitres. The
ends of the second, third, and fourth loops are never coiled into
a regular spiral, but rather form irregularly coiled masses, in many Pelargi,
Accipitres, and in the Psittaci.
We see, then, that the cyclocoelous (mesogyrous or
telogyrous) feature by itself cannot be taken as a characteristic which
indicates the affinity of the larger groups or Orders of Birds,
unless we take the mode of development of these concentric convolutions
into consideration. In fact, the cyclocoelus formation is the highest mode
of stowing in the smallest compass that portion of the gut which had to
be increased in length, the relative length of the mid-gut being dependent
upon the nature and composition of the food. In strictly orthocoelous birds
the increased length of the gut causes the formation of secondary folds
anywhere between the previously existing loops, whereby frequently a very
irregular arrangement of all the convolutions is caused. A similar process
has produced the plagiocoelous
feature (figure f in the diagram
above), which was probably derived from an orthocoelous basis.
The highest and perhaps newest mode of stowing
an increased amount of intestinal length is that in which one of the folds
already existing is lengthened and, owing to its interstitial growth, turns
into a spiral. In this way the other loops will undergo the least
possible disturbance.
|


